作者:Myelin
文章约3000字,阅读约需要4-5分钟
正文
情节记忆 (episodic memory) 是属于长时记忆的一种。被定义为对发生在特定时间和地点的事件的记忆。这种记忆与个人的亲身经历分不开,最大特点是具有情节性,就比如想起自己参加过的一个活动或曾去过的地方。[1]
在我们的日常生活中,我们通常把空间和时间想象成作为经验的独立维度,但我们经常在表达情节记忆时将它们结合起来。如果我问起你的早晨干了什么,你可能会回顾整个早上发生过的事件,因为它是在时间和地点上展开的。发生的地方。这里反映了一种普遍的观点:情节记忆包括将我们对事件的记录嵌入到一个统一的时空背景表述中。[2]
此外,在应用物理学与宇宙学中, “时间”被直截了当地将作为第四维。最著名的是,爱因斯坦的狭义相对论的一个关键组成部分:时间在不同的参照系中会随着速度的变化而扩张。这一观察构成了我们现代 “时空 (spacetime) “概念的基础,即空间和时间维度的统一。[3]
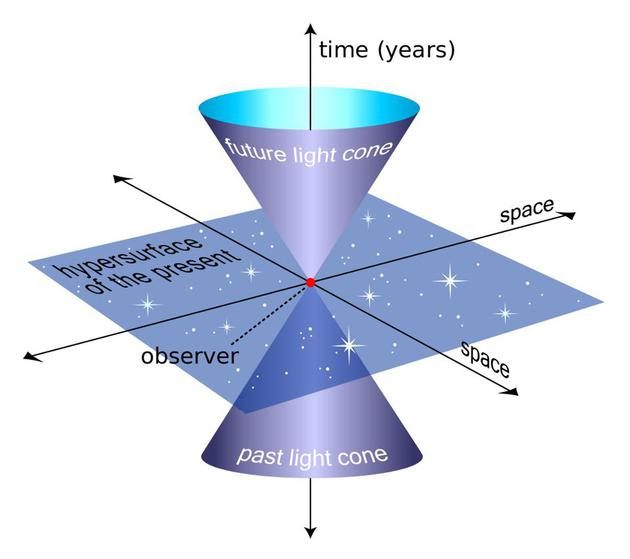
[Fig1. Spacetime,
图片源自:http://www.thephysicsmill.com/2012/12/02/ftl-part-3-general-relativity-shortcuts/]
然鹅,这篇文章将主要介绍大脑是如何处理空间和时间的,重点在于大脑处理空间和时间的途径和机制是反映不同的空间和时间编码?还是反而反映了一个统一的时空表征,在这个表征中,记忆是否直接被定位了下来?
“空间”
长期以来,海马体一直被认为是记忆的关键,也是支持大脑空间和时间表述的关键。这些特征之间的一个潜在联系是:海马体在空间中组织记忆。区分 “什么(what) “和 “哪里(where) “的开创性研究视觉处理的流确定了一个专门的大脑空间认知和行动的专用途径,与支持特定视觉(比如色彩和亮度)的不同途径分开。
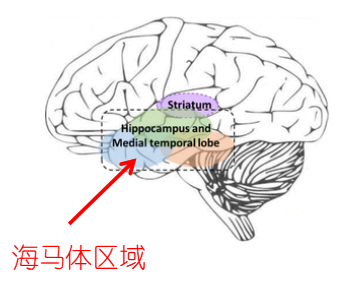
近几十年的研究已经划定了 “哪里 “的细节。“哪里'',也就是空间记忆被编码于连续的皮质区域,从皮质区域(cortex area)延伸到海马旁皮层 (parahippocampal cortex,在灵长类动物中和啮齿类动物中称为脑后皮层)和内嗅皮层 (entorhinal cortex)的内侧部分。[4]

[Fig2.Fig 3.Fig 4 海马体,皮质区与空间记忆,
图片源自:https://www.researchgate.net/profile/Neil-Burgess-2/publication/5580634/figure/fig1/AS:267308312559622@1440742676688/The-hippocampus-and-its-connectionsa-The-hippocampus-lies-in-the-medial-temporal_Q320.jpg]
许多功能成像研究观察到,当受试者回忆起研究特定物品的场景时,海马旁和内侧内皮层会出现选择性激活,从而使研究人员确定这些区域代表了记忆的空间元素。[5]
2005年,May-Britt 和 Edvard Moser发现了网格细胞,这些细胞通过将空间划分为六边形单位,在不同尺度上映射我们的环境。2014年,Moser夫妇与他们在伦敦大学学院的同事和导师John O'Keefe共同获得了诺贝尔生理学或医学奖,因为他们发现了构成大脑定位系统的细胞。
[ Fig.5 嗯, 就比如说这张图片,拿我们的校园举个栗子!教室这个‘空间’就可以被单独用一个六边形表示,每一个边代表一个方位;而食堂这个空间可以用另外一个六边形表示,那么你就可以在上课开小差的时候来规划下课冲向食堂的道路,嘿嘿 这个“道路”就是这里链接六边形示意图里的箭头。]
受Moser夫妇发现空间编码网格细胞的启发,当时卡夫利研究所的博士生Albert Tsao在2007年开始破解谜一样的外侧内视皮层(LEC)中发生的事情。大脑的这一区域紧挨着内侧脑皮层(MEC),他的导师莫泽尔夫妇在那里发现了网格细胞。
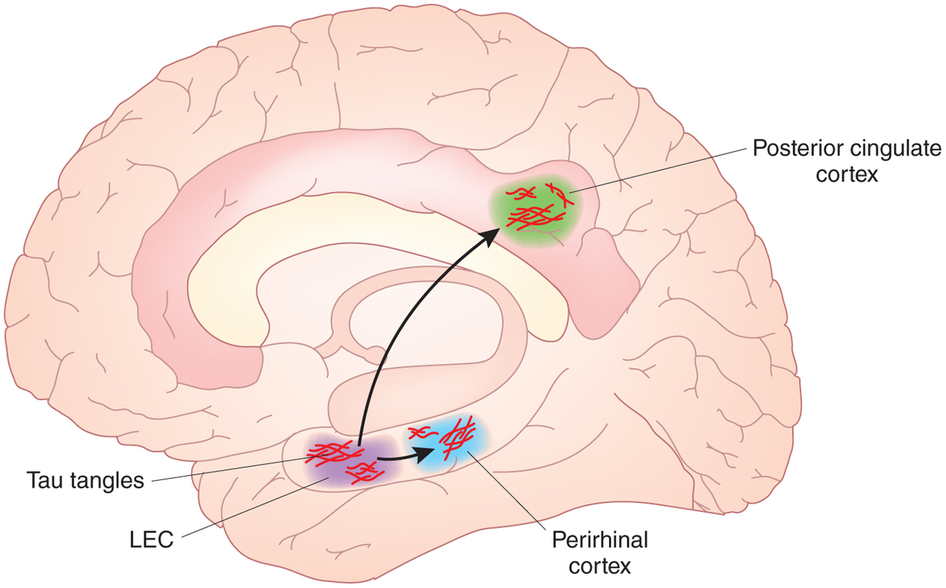
[Fig 6 . LEC,
图片源自:https://i1.wp.com/www.lec.guru/wp-content/uploads/2017/12/Decoding-the-brain.jpg?resize=300%2C227&ssl=1]
今天,我们对我们的大脑处理空间的方式有了相当好的了解,而我们对时间的了解却不那么连贯。时间是一个非平衡的过程。它总是独特和不断变化的,如果这个网络确实在为时间编码,那么信号就必须随着时间的变化而变化,以便将经验记录为独特的记忆。[6]
“时间”
挪威Kavli系统神经科学研究所的研究人员发现了一个脑细胞网络,它在经验和记忆中表达了我们的时间感。神经时钟的运作方式是将我们的经验流组织成一个有序的事件序列。这种活动产生了大脑的主观时间的时钟。因此,经验和经验中的事件序列是主观时间由大脑产生和测量的物质。
就比如说 我们按照时钟时间来协调我们自己的活动。然而,你的大脑并不以你手表上的分钟和小时的标准化单位来感知时间的持续。我们的经验和记忆中的时间特征完全属于一种不同的时间性。[7]
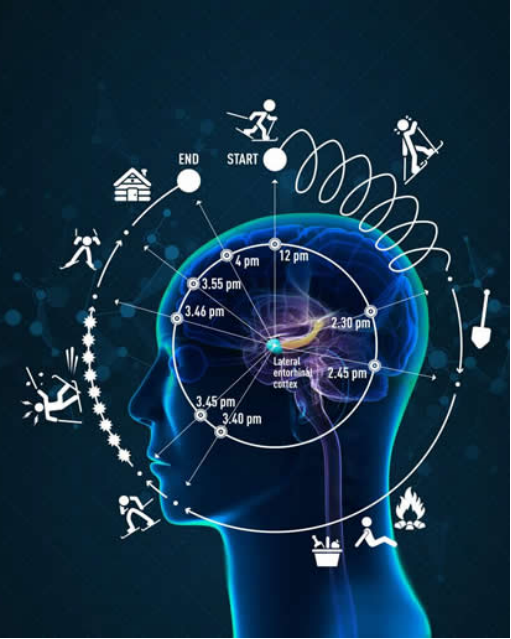
[Fig 7. Time and Brain –
https://norwegianscitechnews.com/wp-content/uploads/2018/08/illustration-1_a-neural-clock-for-timesmall.jpg]
在进化的过程中,包括人类在内的生物体已经发展出多种生物钟来帮助我们掌握时间。大脑的各种计时器的区别不仅在于所测量的时间尺度,还在于神经钟所调整的现象。有些是由外部过程设定的,如昼夜节律钟,它被调谐到日光的上升和下降。这个时钟帮助生物体适应一天的节奏。[8]
其他的计时是由更多内在的现象设定的,比如海马的时间细胞形成了一个多米诺骨牌式的连锁信号,精确追踪时间跨度达10秒。然而,对于大脑用来记录我们的经历和记忆的时间尺度却不多,这些经历和记忆可以持续数秒、数分钟到数小时不等。[7]
2016年,博士生Jørgen Sugar加入了挪威Kavli项目,进行了一组新的实验,以测试LEC网络对偶发时间进行编码的假设。在一个实验中,一只老鼠被引入了一系列广泛的经验和行动选择。它在访问一系列开放空间环境时,可以自由地跑来跑去,调查和追逐巧克力碎片。

[Fig 8 (a). K.Jeffery.
University College London, Topics in Neurobiology]
这个实验中时间信号的唯一性表明,在实验持续的两个小时内,大鼠对时间和事件的时间顺序有非常好的记录,(比如这个耗子它在何时吃到了什么地方的巧克力,看它能不能记得什么时候去哪里接受投喂~)实验人员能够使用来自时间编码网络的信号来准确追踪实验中各种事件发生的时间。
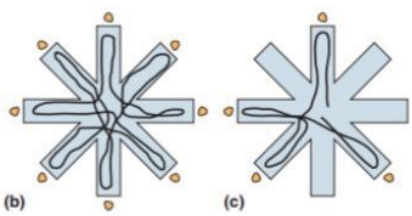
[Fig 8(b) (c )]
在第二个实验中,任务更加结构化,行动的经验和选择范围更窄。大鼠被训练成在一个迷宫中左转或右转时追赶巧克力碎片 [Fig 8 (b) (c)]。通过这项活动,可以观察到时间编码信号从独特的时间序列变成了重复的和部分重叠的模式。
另一方面,在重复性的任务中,时间信号变得更加精确和可预测。这些数据表明,大鼠在每一圈中对时间性的理解很精炼,但在整个实验过程中,从一圈到另一圈以及从开始到结束对时间的理解却很差。
Moser教授认为这项研究表明,通过改变一个人所从事的活动、经验内容,这个人实际上可以改变LEC中时间信号的进程,从而改变人对时间的感知方式。[6,8]
长期以来,时间的结构一直是哲学家和物理学家争议的话题。新发现的大脑对偶发时间的机制能告诉我们什么是我们对时间的感知?我们对时间的感知是像流动的河流一样的线性,还是像车轮或螺旋一样的循环?O‘Keefe 研究的数据表明,这两种情况都是正确的,而且时间编码网络中的信号可以根据经验的不同而采取多种形式。
空间和时间最初是由重叠的大脑网络处理的,并以不同的尺度进行编码,然后空间和时间信号在海马区被整合,形成记忆的空间-时间组织框架。虽然穿越空间和时间的运动在本质上是耦合的,但在某些行为要求下,空间和时间编码可以在海马区不同地观察到,或者由两个维度编码的神经元结合在一起,形成整体的群体表征。
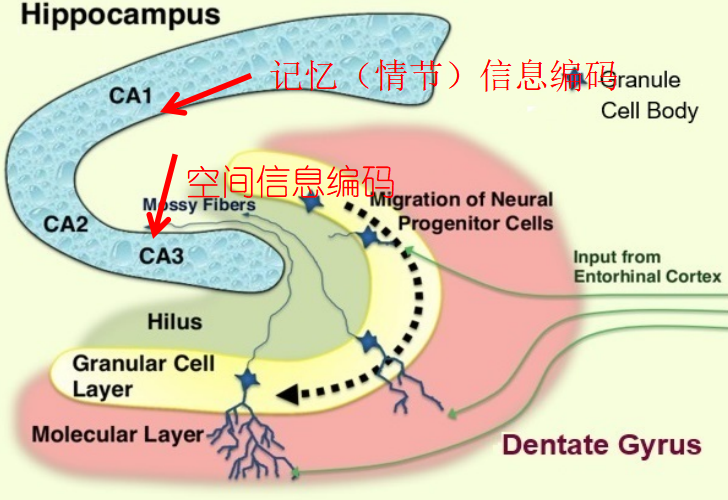
[Fig 9. 海马体CA1,CA3区域及其功能,
图片源自:https://www.geon.us/Memory/images/DentateGyrus.jpg]
不过海马体比较任性(bushi ,海马网络通过对独立编码的空间和时间的混合,与统一的时空坐标表征不同, 选择性将空间和时间表征复用,也就是说时间和空间记忆的混淆也会偶尔发生。
海马网络也映射了广泛的其他连续的经验维度,通过同一群体中神经元元素的混合选择性组织知识。时间背景模型,应用于对不同输入具有高范围的混合选择性的海马网络,表明有一个简单的机制来关联构成空间、时间和其他维度的整合的邻近事件。
呐,这些维度就是抽象的关联空间的特征~
参考文献:
[1] Buzsáki, G. Time, space and memory. Nature 497, 568–569 (2013). https://doi.org/10.1038/497568a
[2] Sarah DuBrow, Lila Davachi, Temporal binding within and across events, Neurobiology of Learning and Memory, Volume 134, Part A , Pages 107-114(2016), https://doi.org/10.1016/j.nlm.2016.07.011.
[3] Brunetti, Romeo, Franceschini, Lorenzo & Moretti, Valter, 2009. Topological features of massive bosons on two-dimensional einstein space-time. Annales Henri Poincaré, 10(6), pp.1027–1073.
[4] Robert E. Clark, Larry R. Squire, Animal model of human memory impairment, Proceedings of the National Academy of Sciences Jun 2013, 110 (Supplement 2) 10365-10370
[5] ALEXANDRA O. CONSTANTINESCU, JILL X. O’REILLY, TIMOTHY E. J. BEHRENS, Organizing conceptual knowledge in humans with a gridlike code, SCIENCE17 JUN 2016 : 1464-1468
[6] Edvard I. Moser, Emilio Kropff and May-Britt Moser ., Place Cells, Grid Cells, and the Brain's Spatial Representation System
Annual Review of Neuroscience.(2008) , Vol. 31:69-89
[7]O’Keefe, J., and Nadel, L. (1978). The Hippocampus as a Cognitive Map(Oxford University Press).
[8]Roberts, B.M., Hsieh, L.-T., and Ranganath, C. (2013). Oscillatory activityduring maintenance of spatial and temporal information in working memory.Neuropsychologia51, 349–357.
图片引用:
Fig. 1: Spacetime, http://www.thephysicsmill.com/2012/12/02/ftl-part-3-general-relativity-shortcuts/
Fig. 2, Fig.3, Fig.4:海马体,皮质区与空间记忆https://www.researchgate.net/profile/Neil-Burgess-2/publication/5580634/figure/fig1/AS:267308312559622@1440742676688/The-hippocampus-and-its-connectionsa-The-hippocampus-lies-in-the-medial-temporal_Q320.jpg
Fig. 6: https://i1.wp.com/www.lec.guru/wp-content/uploads/2017/12/Decoding-the-brain.jpg?resize=300%2C227&ssl=1
Fig. 7: https://norwegianscitechnews.com/wp-content/uploads/2018/08/illustration-1_a-neural-clock-for-timesmall.jpg
Fig.8 (a) (b) (c) : K.Jeffery. University College London, Topics in Neurobiology
Fig 9 : https://www.geon.us/Memory/images/DentateGyrus.jpg
作者:Myelin
未经允许禁止转载或商用
科普平台:https://jq.qq.com/?_wv=1027&k=BCtwKu2W














奖励金币:5
奖励理由:内容饱满(如复制粘贴的使用橡皮擦清除格式,文章使用行间距为2,段落空行或使用段前距,段后距,图片居中,参考文献使用灰色字体加以区分,等丰富排版功能,使其行文,阅读舒适,美观,可增加奖励哦♪)